„”
1. The origin of CRISPR/Cas: from a bacterial immune system to a universal gene editing system
To understand how CRISPR/Cas works as a gene editing system, it is useful to consider its origin as a bacterial immune system interfering with virus or other foreign DNA (reviews: [1, 2]).
Following viral DNA entry into a bacterial cell, bacteriophage DNA and proteins start to control cell metabolism and finally lead to the production of new bacteriophages and cell lysis (lytic cycle; e.g. E. coli T4 phage). Other viruses integrate their DNA temporarily into the host genome before forming new virus particles (lysogenic cycle; e.g. E. coli l phage). As a defense mechanism, bacterial cells have developed CRISPR loci encoding species-specific CRISPR/Cas systems. The best-studied CRISPR locus, that of Streptococcus pyogenes, is shown in Fig. 1. A CRISPR locus encodes the trans-activating CRISPR-RNA (tracrRNA), the Cas operon and the CRISPR repeat spacer array. The eponymous CRISPR repeat spacer array consists of short palindromic repeat sequences which are separated by heterogeneous spacer sequences representing the viral (foreign) DNA. How do the components of the CRISPR locus work together to mediate immunity against viruses? Three distinct steps are involved: i) foreign DNA acquisition, ii) maturation and iii) interference.
When a bacterial cell faces a novel virus DNA, the virus sequence is cut into pieces and fragments are integrated as a novel spacer element into the CRISPR repeat/spacer array (A in Fig. 1; repeat elements = black squares; spacer elements = diamonds in different colors). This DNA acquisition is mediated by the proteins of the Cas operon. Next, maturation takes place. A large RNA is transcribed from the CRISPR repeat/spacer array, the so called pre-CRISPR-RNA (pre-crRNA) (B in Fig. 1). Another component, the pre-trans-activating CRISPR RNA (pre-tracrRNA), is transcribed from the tracrRNA locus (C in Fig. 1). It is complementary to the repeat domains of the pre-crRNA and binds to it via base pairing. Both RNAs form a ternary complex together with the endonuclease Cas9 from the Cas operon (D in Fig. 1). Thereafter, pre-crRNA and pre-tracrRNA are trimmed to their final length by the RNaseIII enzyme (E in Fig. 1). The pre-crRNA is thereby cut into pieces. As a result, virus-specific ternary complexes are formed consisting of the Cas9 protein and the mature tracrRNAs and crRNAs (F in Fig. 1).
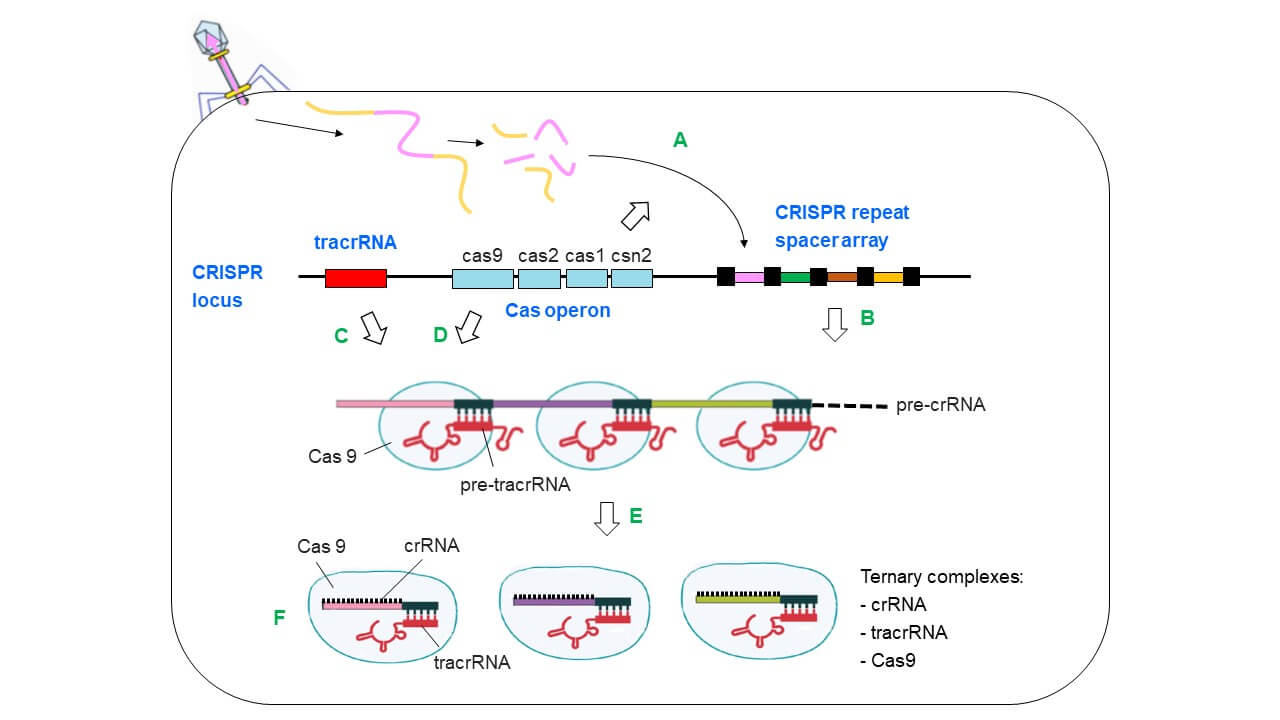
Fig. 1. Foreign DNA acquisition and maturation mediated by the CRISPR locus of S. pyogenes. The individual steps are shown until the virus-specific ternary complexes consisting of Cas9, crRNA, tracrRNA are formed. See the text for details. Adapted and modified from [3]
Interference happens when a virus encounters the cell for a second time (Fig. 2). The ternary complex scans the viral DNA for a Cas-specific trinucleotide sequence, the protospacer adjacent motif (PAM) site. If a PAM site was present and the upstream viral DNA was complimentary to the unpaired crRNA, the ternary complex binds to the target DNA via crRNA/DNA base pairing following unwinding of the target DNA. Cas9 possesses two endonuclease domains, RuvC and HNH which make a double strand break into the invading target DNA thereby destroying it. Of note and in contrast to higher organisms, CRISPR/Cas represents not only an adaptive but also an inherited immune system!

Fig. 2. Interference mediated by the ternary complex of Cas9, crRNA and tracrRNA. Ternary complexes consisting of Cas9, crRNA, tracrRNA make a double strand break into the target DNA once a virus challenges a cell for a second time. See the text for details.
The various bacteria possess different CRISPR loci which fall into different classes and sub-types depending mainly on the organization of the nuclease portion. According to the classification shown in Fig. 3, the CRISPR locus of S. pyogenes shown above represents a class 2, type II system.

Fig. 3. Stucture and classification of various CRISPR systems. The various bacteria possess different CRISPR/Cas systems. In particular, the organization of the nuclease function (red) is highly variable. Adapted and modified from [4]
The history of CRISPR/Cas started in the early nineties. The fact that the spacer/repeat domain contains viral DNA was described in 1993 for the archaebacterium Haloferax mediterranei [5]. After that, it took almost 20 years to describe all components of a CRISPR locus and to understand their interplay within a bacterial immune system. After description of the last component of the system, namely the tracrRNA [6]), and the species interchangeability of the system in 2011 [7], it was speculated that CRISPR/Cas could serve as a novel programmable, RNA-based gene editing system. Such an application was shown in vitro in 2012 by two groups almost in parallel [8] [3] - in mammalian cells only a few months later [9] [10].
2. CRISPR/Cas as an universal gene editing system: the basis of knock-outs and knock-ins.
How does gene editing with CRISPR/Cas work? To target a gene in cells, a Cas protein is needed (most frequently Cas9) which is complexed with a single guide RNA (sgRNA). The sgRNA represents a fusion of the tracrRNA and a user-designed crRNA portion which is complimentary to an exon of the target gene. Suitable sgRNAs are found by bioinformatics approaches. To express both Cas9 and the specific sgRNA, usually the same vector plasmid is used. Plasmid delivery into cells is either mediated by lipofection or viral transfer. Electroporation using preformed, purified Cas9/sgRNA complexes is also frequently used.
If a gene knock-out in eukaryotic cells is intended, the complex binds via sgRNA base pairing to the target sequence. Cas9 will make a double strand break into the target gene and the often faulty repair causes small insertions or deletions (indels). This so called non-homologous end joining (NHEJ) is used to create knock-outs of genes (KOs). On the other hand, double strand breaks made by Cas9 are known to stimulate homologous recombination. By presenting specific repair templates, it is also possible to introduce specific modifications into the target. This process is called homology-directed repair (HDR) and is used to create knock-ins (KIs). A summary of the processes leading to KOs and KIs is shown in Fig. 4. We invite you to do perform your intended KOs and KIs in cooperation with our facility. In case other applications are needed (see below) we could discuss them, too.
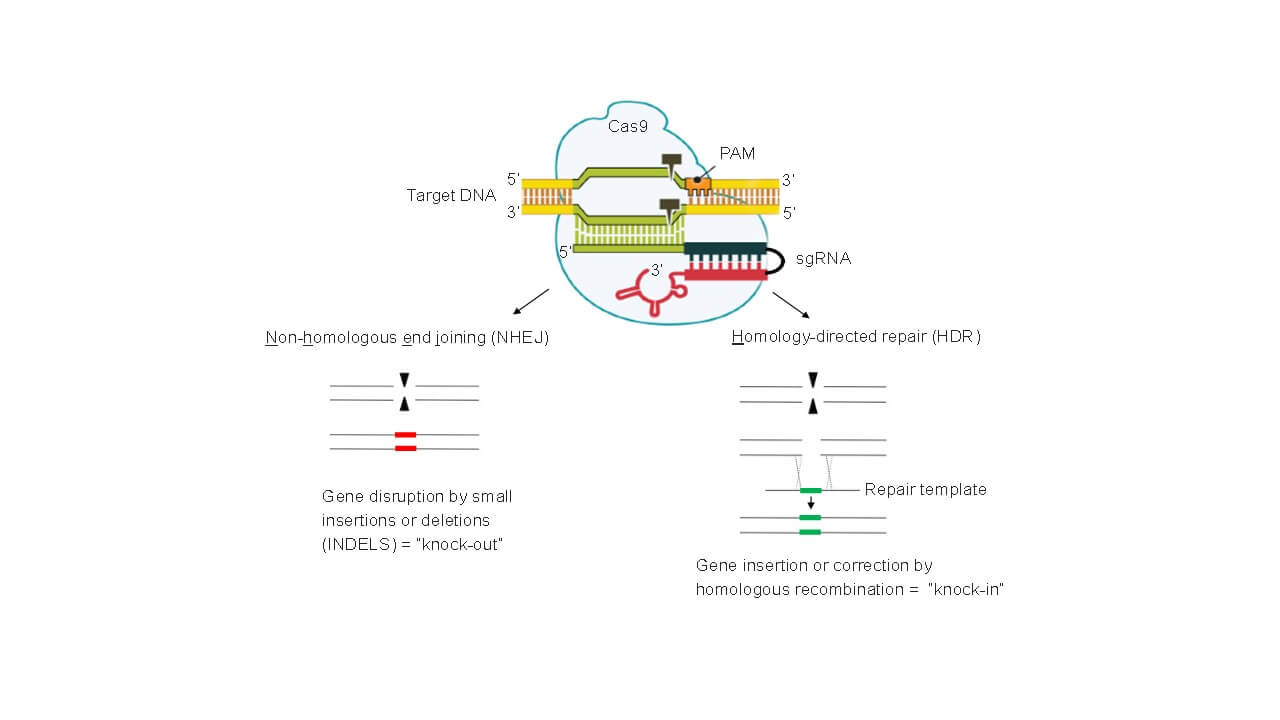
Fig. 4. CRISPR/Cas: creating KOs and KIs. Non homologous end-joining can be used to create indels in the target DNA (KOs), an additional repair template can be used to produce KIs by homology-directed repair.
3. Examples for other applications of CRISPR/Cas
Within the last six years, the original CRISPR/Cas9 gene editing system for KOs and KIs was improved substantially in terms of PAM site use and Cas/sgRNA delivery into cells. Even a non-cloning procedure with purified Cas9, synthetic sgRNA, and electroporation is available and works efficiently. Inactivation of either one of the two nuclease domains of Cas9 leads to a single-strand breaking Cas9 (nickase, nCas9). Tandem use of two parallel nCas9 proteins was shown to improve targeting specificity.
Of note, not only the original CRISPR/Cas system for KOs and KIs was improved. The number of additional applications has also grown substantially. If both nuclease domains are inactivated, a Cas protein results which is unable to cleave DNA but still binds to the specific target sites (dCas, Fig. 5). Such dCas variants can be fused for example with activators or repressors of gene expression or with epigenetic markers or fluorescent proteins. The latter may be used as novel diagnostic tools to detect viruses or infectious bacteria in the future.

Fig. 5. Examples for applications using dCas9. Target DNA binding but non-cutting dCas9 can be fused with gene activation or gene repression domains or with epigenetic modifiers. Fusions with fluorescent proteins may help to detect a specific target DNA.
4. Base editors for “second generation CRISPR/Cas genome editing”?
The possibility to fuse dCas9 with epigenetic markers raised the question whether the nuclease may be fused with base editors, too. The successful fusion of cytidine and adenine desaminases to dCas9 turned an epigenetic marker again into a genetic marker which works without cutting DNA. These enzymes induce specific nearby base conversions (C→T; A→G) and stop codons were introduced successfully by this technique into target genes. Base editors may pave the way to site-directed mutagenesis of the genome, the so called second generation CRISPR/Cas genome editing.
5. Problems and drawbacks of CRISPR/Cas. There are still some drawbacks of CRISPR/Cas which should be taken into account. You may ask why we don’t have a CRISPR/Cas system which could fight against viruses and could be inherited. The answer is most likely the CRISPR/Cas is not very specific and may cause a lot of trouble in genome which is a problem for higher organisms but not for bacteria with their high reproduction rates. The most obvious drawbacks of CRISPR/Cas are off-target effects resulting from the fact the target-specific moiety in the sgRNA is only 20 bases long. Off-target effects are thus common during gene editing, but in principle predictable by bioinformatics. It should be taken into account, however, that even the best sgRNA design may lead to off-target effects. You will thus get potential off-target sites from us and you should in particular look whether they would affect your pathway of interest. The sgRNAs should be re-disigned in this case. Alternatively, it should be determined afterwards experimentally that these off-targets did not happen.
References
1. Lander ES. The Heroes of CRISPR. Cell. 2016;164(1-2):18-28.
2. Thurtle-Schmidt DM, Lo TW. Molecular biology at the cutting edge: A review on CRISPR/CAS9 gene editing for undergraduates. Biochemistry and molecular biology education : a bimonthly publication of the International Union of Biochemistry and Molecular Biology. 2018;46(2):195-205.
3. Doudna JA, Charpentier E. Genome editing. The new frontier of genome engineering with CRISPR-Cas9. Science. 2014;346(6213):1258096.
4. Mohanraju P, Makarova KS, Zetsche B, Zhang F, Koonin EV, van der Oost J. Diverse evolutionary roots and mechanistic variations of the CRISPR-Cas systems. Science. 2016;353(6299):aad5147.
5. Mojica FJ, Juez G, Rodriguez-Valera F. Transcription at different salinities of Haloferax mediterranei sequences adjacent to partially modified PstI sites. Molecular microbiology. 1993;9(3):613-21.
6. Deltcheva E, Chylinski K, Sharma CM, Gonzales K, Chao Y, Pirzada ZA, et al. CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III. Nature. 2011;471(7340):602-7.
7. Sapranauskas R, Gasiunas G, Fremaux C, Barrangou R, Horvath P, Siksnys V. The Streptococcus thermophilus CRISPR/Cas system provides immunity in Escherichia coli. Nucleic acids research. 2011;39(21):9275-82.
8. Gasiunas G, Barrangou R, Horvath P, Siksnys V. Cas9-crRNA ribonucleoprotein complex mediates specific DNA cleavage for adaptive immunity in bacteria. Proc Natl Acad Sci U S A. 2012;109(39):E2579-86.
9. Cong L, Ran FA, Cox D, Lin S, Barretto R, Habib N, et al. Multiplex genome engineering using CRISPR/Cas systems. Science. 2013;339(6121):819-23.
10. Mali P, Yang L, Esvelt KM, Aach J, Guell M, DiCarlo JE, et al. RNA-guided human genome engineering via Cas9. Science. 2013;339(6121):823-6.
The Leibniz-Forschungsinstitut für Molekulare Pharmakologie (FMP) is part of the Forschungsverbund Berlin e.V. (FVB), which legally represents seven non-university research institutes - members of the Leibniz Association - in Berlin.
Leibniz-Forschungsinstitut für Molekulare Pharmakologie im Forschungsverbund Berlin e.V. (FMP)
Campus Berlin-Buch
Robert-Roessle-Str. 10,
13125 Berlin, Germany